430 млн. л. н. на планету упал метеорит. Резкое понижение уровня мирового океана.
Оледенение Гондваны окончилось в самом начале Силурийского Периода, и рубеж раннего и позднего палеозоя вновь характеризуется развитием одинаково высоких температур, как в низких, так и в высоких широтах.
ЭПОХА РЫБ И АМФИБИЙ
Развитие из кожных складок плавников повысило маневренность движения в воде, парные грудные и брюшные плавники приобрели хрящевой или костный скелет-сформировались пояса конечностей.
Хрящевые рыбы - класс рыбообразных позвоночных, осевой скелет которых целиком состоит из хряща. Силур-ныне.
ПОЗВОНОЧНЫЕ
Цефаласпис - вымерший род костнопанцирных остракодерм, примитивных морских бесчелюстных позвоночных. Жил в верхнем силуре — среднем девоне в водах Лавруссии (современные Северная Америка и северо-западная Европа). Цефаласпис — один из первых открытых представителей остракодерм. Этот род описал Луи Агассис в 1835 г., отнеся к нему 4 вида. Два из них позже оказались представителями разнощитковых, третий — идентичным по крайней мере одному из первых двух, a четвёртый, Cephalaspis lyelli, и сейчас служит типовым видом рода. Первые исследователи считали цефаласписа, как и других известных тогда остракодерм, костной рыбой. К бесчелюстным их отнёс в 1889 г. Эдвард Коп, хотя окончательно отсутствие у цефаласписа челюстей доказал Эрик Стеншё только в 1920-х гг. Со времён Агассиса к этому роду относили очень много видов (зачастую плохо изученных), которые напоминали C. lyelli формой головного щита (состоянием на 2004 год известно 125 видов, когда-либо побывавших в этом роде). Частично эта ситуация объясняется недостаточно подробным описанием типового экземпляра. Впоследствии большинство этих видов отнесли к другим родам, а многие — и к другим семействам. В одной из последних систем род Cephalaspis включает 4 вида (C. lyelli, C. cradleyensis, C. producta и C. sollasi), и не исключается, что более детальные исследования ограничат его объём типовым видом. Характерные признаки рода — довольно длинная преорбитальная часть головного щита, параболическая форма края этого щита и узкое срединное поле мелких костных пластинок на нём. Цефаласпис считается относительно продвинутым родом костнопанцирных. Животные этого рода достигали 60 см. в длину и спереди были покрыты массивным панцирем, который служил им защитой от хищников, но заметно снижал подвижность. Зато у них была хорошо развита мускулатура задней (гибкой) части тела и, в отличие от большинства остракодерм, грудные плавники. Поэтому подвижность и манёвренность цефаласписа считается относительно большой, хотя, как и у всех остракодерм, его приспособления к плаванию были намного менее эффективны, чем у рыб и его образ жизни был, по-видимому, в основном придонным. Для цефаласписа и других сходных с ним костнопанцирных предполагается способность к довольно продолжительному планированию в воде. Это позволяла его уплощённая брюшная поверхность, дополнительно увеличенная за счёт «рогов» (выростов по бокам головы, направленных назад). «Рога», кроме того, выполняли роль стабилизаторов во время плавания. Цефаласписы были лишены челюстей и, вероятно, питались донным детритом на коралловых рифах. В свою очередь, служили пищей крупным хищникам, таким как ракоскорпионы и панцирные рыбы. У цефаласписов была развита система обнаружения опасности, особые сенсоры на его кожи улавливают в воде малейшие вибрации (ЧУВСТВО ОСЯЗАНИЯ). Но защитные приспособления не позволяют цефаласпису долго плыть с большой скоростью ему часто приходится отдыхать. Врагами цефаласписов были членистоногие.
Модель представителя ракоскорпионов, демонстрируемая в Национальном музее естественной истории в Вашингтоне.
Ракоскорпионы, или морские скорпионы, или эвриптериды (лат. Eurypterida) — ископаемый отряд членистоногих из класса Меростомовые подтипа (по другой систематике — надкласса) хелицеровых (Chelicerata). Отдельные представители достигали 2 метров в длину, однако характерные размеры большинства видов не превышали 20 см. Ранние формы обитали на мелководье в морях. На примере эволюции ракоскорпионов хорошо прослеживается переход от водного образа жизни к сухопутному. У этих созданий есть жабры, у них есть жало размером с электрическую лампочку. Это главные хищники Силурийсих морей (Силур-ранний Девон). Охотились они на морском дне на цифоласписов.
Окаменелость ракоскорпиона Eurypterus remipes.
С наступление брачного периода цефаласписы устремляются туда где до них не доберётся ракоскорпион в пресную заводь удалённую от моря.
Суша в то время представляла из себя бесплодные просторы покрытые горными породами. Воздух содержал мало кислорода и в 300 раз больше углекислого газа.
Цефаласпис пробирается к верховью реки, удаляясь от моря. Он проделает долгий путь прежде чем окажется в месте размножения, где сам он появился на свет. Их укреплённые черепа защищают жизненно важное орудие - первый сложный головной мозг. Он намного более развит чем у их соперников ракоскорпионов, у которых совсем нет памяти.
У ракоскорпионов помимо жабер были примитивные лёгкие, состоящие из сотен тонких слоёв ткани. Они имея приспособление позволяющее максимально использовать малое количество кислорода, а также панцирь защищающий от Солнца, патрулировали берег подбирая всё что выбрасывало море. Ослабленным после долгого путешествия рыбам придётся переправляться из реки в пруд, преодолевая высокие пороги. На обессиленных цифоласписов на порогах нападают ракоскорпионы. Уцелевшие рыбы, переправившись в озеро, начинают метать икру, потом они опять возвращаются в море.
Гондванаскорпио
Гондванаскорпио - эвриптерид, найденный в ЮАР и живший 350 млн лет назад. Палеонтологи, ведущие современные раскопки на территории ЮАР, обнаружили недалеко от южноафриканского города Гремстаун окаменевший отпечаток древнего скорпиона, который обитал на Земле около 350 млн лет назад, в самом начале силурийского геологического периода. По мнению исследователей — это самый древней из известных науке обитатель континента Гондвана, когда-то существовавшего на Земле. Поскольку в то время другой живности кроме насекомых не было, то скорпионы, пауки и ящерицы питались только ими. Найденный вид получил название Gondwanascorpio emzantsiensis. Ученые надеются, что находка даст возможность больше узнать о жизни животного мира на Гондване до момента его раскола.
Джекелоптерус
Джекелоптерус (Jaekelopterus) - эвриптерид пресных водоёмов Германии. Жил 400 м.л.н. Длина до 2,5 м (предположительно). Длина клешни 46 см. Достиг таких размеров из-за большого содержания кислорода, относился к водным животным.
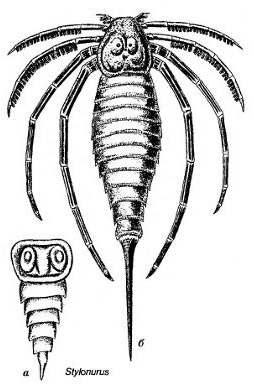
Стилонурус
Стилонурус - вымерший вид эвриптерид длиной 3 м. Стилонурус сочетал длинный тельсон и сужающийся абдомен с относительно маленькой и очень четкой просомой, на которой сложные глаза придвинулись почти к центру. Хелицеры (первая пара) были короткими. Следующие конечности были ходильными, из них первая пара конечностей была короткой и шипастой, вторая и третья пары были длинными с шипами, расположенными, как зубцы гребенки, а две последние пары были ещё длиннее и несли продольные длинные гребни. Эти последние конечности выглядят совсем непохожими на те лопатовидные и весловидные конечности остальных эвриптерид, оканчивающихся острыми загнутыми коготками. Стилонурус несомненно ходил на этих восьми конечностях, а шипастые из них мог использовать для удержания пищи, так как хелицеры были короткими.
Трилобиты (членистоногие) в длину достигли до 75 см.
Ортоцерас
Ортоцерас — род вымерших головоногих, дальний родственник кальмаров и каракатицы, только очень длинный. Ортоцерасы как и ортоконы имели длинные раковины, только были они гораздо меньше своих родственников. Длина обычного ортоцерата всего около 2 метров. Ортоцераты обитали в Ордовикский период и вымерли в конце Триасового периода. Ортоцерасов относят к отряду Ортоцерид (пряморогов). Подобно наутилусам и аммонитам ортоцераты обладали большой, хорошо развитой раковиной, но в отличие от них, у ортоцерат раковина была не спиралеобразной, а конусообразной. Раковины ортоцерат внутри были заполнены множеством камер, заполненных воздухом или другими газами. Последняя камера была самой обширной. При опасности ортоцерат мог прятать в ней голову и щупальца. На голове ортоцерата имелись большие глаза и множество щупалец. Как и у современных наутилусов, у ордтоцерат на щупальцах отсутствовали присоски. Щупальцами они захватывали свою добычу (различных некрупных животных) и подтягивали ее ко рту, где располагался крепкий клюв. Массивная прямая раковина, по-видимому, создавала ортоцератам определённые трудности при плавании и особенно при маневрировании. В силу этого они в основном обитали вблизи морского дна, медленно перемещаясь в поисках добычи. Окаменелости ортоцерат можно найти в Марокко, Скандинавии, в Альпах, и в Айове (США).
ПОЗДНИЙ СИЛУР
РАСТЕНИЯ
ХАРОВЫЕ - тип, объединяющий самые высокоорганизованные водоросли; родственны, с одной стороны, зелёным водорослям, а с другой - высшим растениям. Относительно крупные растения с дифференцированным слоевищем и сложно устроенными органами размножения. Силур-ныне.
В конце Силура на Земле происходят интенсивные горообразовательные процессы, приведшие к возникновению Скандинавских гор, гор Тянь-Шань, Саян, а также к обмелению и исчезновению многих морей. В результате, некоторые водоросли (сходные с современными харовыми водорослями) выходят на сушу, и заселяют литорали и супралиторали, что стало возможным благодаря деятельности бактерий и цианобактерий, образовавших на поверхности суши почвенный субстрат. Так возникают первые высшие растения — риниофиты. Особенность риниофитов заключается в появлении тканей и их дифференцировки на покровные, механические, проводящие и фотосинтезирующие. Это было спровоцировано резким отличием воздушной среды от водной. В частности: - повышенной солнечной радиацией, для защиты от которой у первых наземных растений должен был выделяться и откладываться на поверхности кутин, что и было первым этапом формирования покровных тканей (эпидермы); - откладывание кутина делает невозможным поглощение влаги всей площадью (как у водорослей), что приводит к изменению функции ризоидов, которые теперь не только прикрепляют организм к субстрату, но и поглощают из него воду; - разделение на подземную и надземную части спровоцировало необходимость доставки минеральных веществ, воды и продуктов фотосинтеза по всему организму реализованную появившимися проводящими тканями — ксилемой и флоэмой; - отсутствие выталкивающей силы воды и соответственно невозможность плавать, в ходе конкуренции видов за солнечный свет, привело к появлению механических тканей с целью «приподняться» над соседями, ещё одним фактором было улучшенное освещение активизировавшее процесс фотосинтеза и приведшее к избытку углерода, что и позволило образоваться механическим тканям; - в ходе всех вышеперечисленных ароморфозов фотосинтезирующие клетки выделяются в отдельную ткань.
Древнейшее известное наземное растение — куксония. Куксония обнаружена в 1937 г. в силурийских песчаниках Шотландии (возраст порядка 415 млн. лет). Дальнейшая эволюция высших растений разделилась на две линии: гаметофитную (моховидные) и спорофитную (сосудистые растения). Первые голосеменные растения появляются в начале Мезозоя (примерно 220 млн. лет назад). Первые покрытосеменные (цветковые) возникают в юрском периоде.
Первоначально существовала (на суше и во временных водоёмах) группа зелёных водорослей ("микроскопические харовые"), от которой в Силуре произошли две близкородственные группы: "настоящие" харовые, заселившие континентальные водоёмы, и высшие растения, начавшие осваивать сушу и лишь спустя некоторое время появляющиеся в прибрежных местообитаниях.
ВЫСШИЕ РАСТЕНИЯ - многоклеточные наземные или вторичноводные растения, тело которых имеет сложно дифференцированные системы органов и тканей. К ним относятся сосудистые растения и мохообразные. Силур-ныне.
В этой обжигающей среде появились самые первые сосудистые растения- риниофиты (тип споровых растений, объединяющий самые примитивные из сосудистых растений-травянистые формы с дихотомическим ветвлением); вплоть до раннего Девона они были представлены крайне однообразными остатками единственного рода Куксония (простейший и архаичный из сосудистых. Силур-Девон) - имеет уникальную стратегию выживания, это первое растение чьи побеги стали тянуться вверх, чтобы получить дополнительный свет для роста. Это в итоге приведёт к возникновению лесов.
Так на побережьях, периодически заливаемых водой, появились первые наземные примитивные сосудистые растения. Они дали начало всем Высшим Растениям (споровым, хвощам, папоротниковым, плаунам).
Появление сосудистых растений с их жёсткими вертикальными осями вызвало целый каскад экосистемных новаций, изменивших облик всей биосферы:
1. Фотосинтезирующие структуры стали располагаться в трёхмерном пространстве, а не на плоскости (как это было до сих пор-в период господства водорослевых корок и лишайников). Это резко увеличило интенсивность образования органического вещества и, следовательно, суммарную продуктивность биосферы.
2. Вертикальное расположение стволов сделало растения более устойчивыми к занесению смываемым мелкозёмом. Это уменьшило безвозвратные потери экосистемой неокисленного углерод а (в виде органики), т.е. произошло совершенствование углеродного цикла.
3. Вертикальные стволы наземных растений должны быть достаточно жёсткими (по сравнению с водными макрофитами) для обеспечения этой жёсткости возникла новая ткань - древесина, которая после гибели растения разлагается относительно медленно. Углеродный цикл экосистемы обретает дополнительное резервное депо и стабилизируется.
4. Появление постоянно существующего запаса трудноразложимой органики (сконцентрированного в основном в почве) ведёт к радикальной перестройке пищевых цепей. С этого времени большая часть вещества и энергии оборачивается через детритные, а не через пастбищные цепи.
5. Для разложения трудноусвояемых веществ, из которых состоит древесина (целлюлозы и лигнина), потребовались новые типы разрушителей мёртвой органики. С этого времени на суше роль основных деструкторов переходит от бактерий к грибам.
6. Для поддержания ствола в вертикальном положении (в условиях действия силы тяжести и ветров) возникла развитая корневая система.
У панцирных бесчелюстных остракодерм (цефаласпис) из жаберных дуг развиваются подвижные челюсти (ротовой аппарат хватательного типа). Питание с активным захватом добычи способствовало развитию нервной системы, перестройке всей организации рыб.
Появились челюстноротые - эволюционная ветвь (инфратип), включающая всех позвоночных (кроме бесчелюстных), т.е. рыб и тетрапод. Силур-ныне.
Плакодермы, панцирные рыбы - подкласс архаичных хрящевых рыб; голова и передняя часть тела у них были покрыты щитками. Конец Силура-Девон.
Артродиры (лат. Arthrodira, от др.-греч. ἀρθρο- +δειρή «сочленённая шея»), членистошейные — отряд вымерших рыб класса Панцирные рыбы (Placodermi). Известны с силура по верхний девон. Полная длина этих рыб неизвестна, так как найдены лишь фрагменты экзоскелета головы, плечевого пояса и позвоночника. Приводится лишь предположительная длина этих рыб, рассчитанная (исходя из размера их головы) по пропорциям современных рыб. Артродиры (в большинстве своём) — хищники-ихтиофаги (особенно крупные виды, обладавшие мощными челюстями с огромной силой сжатия); исключение составляет титанихтис, который был планктофагом — его челюсти были достаточно слабо развитыми. Особенностью строения черепа многих артродир являлось то, что верхняя часть головы могла открываться под равным углом с нижней челюстью наподобие кусачек; это, скорее всего, давало им возможность открывать рот с большой скоростью, тем самым создавая область разрежения (низкого давления) в ротовой полости. В результате создавался поток воды, всасывающий пищевой объект (жертву) прямо в пасть рыбы (это означает, что артродиры скорее были засадными хищниками, чем активными пелагическими преследователями). Экзоскелет головы и плечевого пояса состоял из парных и непарных пластин, сросшихся друг с другом. Каждый геологический период характеризовался своим набором видов артродир с присущими только им чертами строения. Так, более архаичные из артродир — арктолепиды — были распространены в верхнем силуре — нижнем девоне. Для среднего девона характерны представители инфраотряда Coccosteina, у которых наблюдается более прогрессивное строение панциря (увеличивается высота тела, челюсти становятся более сильными). Для верхнего девона обычны рахиостеиды.
Паукообразные - класс наземных членистоногих из подтипа хелицеровых. Силур-ныне.
Членистоногие сделали первые шаги из моря на сушу. На сушу выходят членистоногие растительноядные двупарноногие многоножки (собирательное название трачейнодышащих членистоногих с червообразным телом) и скорпионы (отряд класса паукообразных, связанный прямым родством с ракоскорпионами. Силур-ныне.), появляются тригонотарбы ("панцирные пауки"- отряд класса паукообразных, представители которого первыми заселили сушу. Силур-Карбон.).
Многоножки - надкласс, объединяющий четыре класса наземных членистоногих (симфил, губоногих, двупарноногих и пауропод, последних обычно объединяют в одну группу). Самые древние ископаемые представители надкласса многоножек (Pneumodesmus newmani из класса двупарноногих) были обнаружены в слоях датируемых поздним Силурским периодом (возраст около 428 миллионов лет).
Тригонаторпиды - отряд вымерших паукообразных, одни из первых животных, вышедших на сушу (Силур-Карбон). Первый вид отряда был найден в Великобритании в 1837 году. Ископаемые остатки этих животных находят также в Аргентине, в Южной Америке, в Европе. Тригонотарбиды обитали 419—290 млн. лет назад, с конца Силура до начала Пермского периода. Это самые древние известные сухопутные членистоногие. Всего открыто около 70 видов тригонотарб. Это были хищные членистоногие, питавшиеся другими членистоногими. Очевидно, они охотились на земле. Тригонотарбиды были найдены у самых корней растений, где, возможно, скрывались, ожидая добычу. Как показали палеонтологические находки, такие виды тригонотарб, как Aphantomartus pustulatus, обитавшей 300 миллионов лет назад, использовали паутину (ранее же считалось, что Ттригонотарбы не преуспели в создании паутинопрядильных органов, которые, очевидно, были предпосылкой к эволюционному успеху пауков, но в последние годы найдена, по крайней мере, одна окаменелость, у которой есть microtubercles, родственный используемым пауками, чтобы направлять и управлять их паутиной). Возможно, животное могло использовать паутину для ловли добычи. Однако, тригонотарбы не имели прядильных желез на опистосоме. Возможно, хелицеры тригонотарб были не ядовиты (так как не найдено желез яда хелицеры). Однако, они были очень сильными. Внешне тригонотарбиды напоминают пауков. Тело тригонотарб расчленялось на головогрудь и брюшко. Имелся прочный членистый панцирь (видимо, наследие панцирных хелицеровых, как мечехвосты и ракоскорпионы). Длина тела этих животных составляла от нескольких миллиметров до 5 см. Животные имели пять-шесть пар ног. На некоторых конечностях были коготки, помогавшие при охоте и удержании добычи. Лёгочное дыхание. Органы чувств — глаза, волоски. Тригонотарбиды наиболее родственны современным рицинулеям. Тригонотарбы вымерли уже в Каменноугольном периоде.
ЛАВРУССИЯ
408 млн. л. н. резкое понижение уровня мирового океана.
408 млн. л. н. тектомагматические сдвиги КАЛЕДОНСКОЙ Эпохи.
Началось замыкание океанских бассейнов и постепенное сближение материков. Гондвана медленно перемещалась в юго-западном направлении и соединилась с С. Американской платформой.
В северном полушарии возник крупный материк - ЛАВРУССИЯ (С. Америка, В. Европа). Произошло закрытие океана ЯПЕТУС (ПалеоАтлантический).
Произошли крупные горообразовательные процессы. Образуются высочайшие протяжённые горно-складчатые сооружения-Аппалачи, Ц. Казахстанские, Алтайские, З. и В. Саяны, горы Монголии, В. Австралии, Антарктиды.
События СИЛУРА:
- Расчленение тела на отделы: голова, туловище, хвост (панцирные, щитковые).
- Выход на сушу растений, дифференциация тела на ткани.
Появление сосудистых растений с их жёсткими вертикальными осями вызвало целый каскад экосистемных новаций, изменивших облик всей биосферы.
1) Фотосинтезирующие структуры стали располагаться в трёхмерном пространстве, а не на плоскости (в период господства водорослевых корок и лишайников). Это резко увеличило интенсивность образования органического вещества.
2) Вертикальное расположение стволов сделало растения более устойчивыми к занесению смываемым мелкозёмом. Это уменьшило безвозвратные потери экосистемой неокисленного углерода (в виде органики), т.е. произошло совершенствование углеродного цикла.
 ЕДИНСТВО
ЕДИНСТВО