ПРИМАТЫ
ПРИМАТЫ - отряд млекопитающих, к которому относятся лемуры, обезьяны и люди.
Предками приматов и других плацентарных млекопитающих, были древнейшие насекомоядные - эндотерии, остатки одного из представителей которых были найдены в позднеюрских и раннемеловых отложениях в горах Лаоса, Северо-Восточном Китае. Эндотерии сформировались в одной из частей Годваны, скорее всего, в лемурийской её части, ныне занятой Индийским океаном.
В качестве характерного представителя этой группы, предковой формы для приматов можно указать
залямбдалестеса, неполный череп, которого был найден в слоях
Позднего Мела Монголии. Эти предки тупай вели полудревесный образ жизни. Хвост был длинный, конечности сравнительно короткие, пятипалые с когтями; в запястье - свободные центральная, полулунная и ладьевидная косточки и предплюсне-таранная кость с широким блоком; передняя конечность с развивающейся способностью хватания. Голова относительно крупная, мозг низко - организованный с крупными обонятельными долями (макросоматичность), но уже с признаками начинающегося развития зрительной области. Мордочка длинная, ноздри на конце мордочки. Полость глазницы широко соединилась с височной ямкой.
До распространения покрытосеменных (до Мела) растений ландшафт южных материков Гондваны представлял собой чередование открытых пространств и разреженных лесов, большинство древних насекомоядных были наземными животными. Лишь в конце Мезозоя, когда возникли огромные массивы лесов покрытосеменных, часть насекомоядных, не успевшая специализироваться в поисках и погоне за насекомыми, постепенно стала приспосабливаться к древесному образу жизни. В связи с новым образом жизни на кустарниках и деревьях тело этих насекомоядных должно было претерпеть значительные изменения. В поисках пищи, в погоне за насекомыми им приходилось передвигаться не только по стволам и толстым веткам деревьев, но и по более тонким ветвям, перескакивать с ветки на ветку, с дерева на дерево, лазать по лианам и воздушным растениях (эпифитам). Поэтому, прежде всего изменилось строение конечностей этих насекомоядных.
Жизнь на деревьях сказалась на относительном значении органов чувств этих животных. Примитивное наземное млекопитающее руководствуется в своём поведении преимущественно обонятельными ощущениями. Орган обоняния играет первую роль в жизни такого животного, а обонятельная доля полушарий мозга - наиболее крупная и прогрессивная. С преимущественным существованием на дереве и с отрывом от земли положение меняется; на деревьях восприятие животным запахов теряет значение. Здесь гораздо важнее слух и точный глазомер. У древесных животных прогрессивно развиваются соответствующие органы чувств. Для приматов с их хватательными конечностями, с их легкоподвижными пальцами большую важность приобретают осязательные ощущения, особенно остро воспринимаемые при посредстве чувствительных голых участков кожи, расположенных на конечностях. Концы пальцев приматов в связи с этим расширены; они защищены плоскими ногтями (вместо когтей большинства других млекопитающих) и на их коже, соприкасающейся с окружающими предметами, находятся "тактильные узоры" - ряды тонких валиков, расположенных дугами, петлями, кругами, овалами и т.д.
Усиленное развитие органа зрения и осязания сопровождалось у приматов упадком роли обоняния. Соответственно с этим у них уменьшено число носовых раковин и весь носовой отдел черепа.
Древнейшие приматы возникли в Азии. Они в Меловом периоде распались на две основные группы: протолемуроидов (предков тупай и лемуров) и прототарзиоидов (предки долгопятов). Из Азии они расселились по другим материкам Старого Света и перебрались в Северную Америку.
Тупайеобразные:
Семейство Тупайи (4 рода, 17 видов);
Семейство пёстрохвостые Тупайи (1 вид).
Лемурообразные:
Семейство Адапиды – как лемуры;
Семейство Лемуры (6 родов, 16 видов);
Семейство Лори (4 рода, 5 видов);
Семейство Руконожки (1 вид);
Семейство Галаго (2 рода, 6 видов);
Семейство Индри (3 рода, 4 вида).
Долгопятообразные:
Семейство Омомиды – предки долгопятов;
Семейство Долгопяты (1 род, 3 вида).
Некоторые насекомоядные приспособились к передвижению прыжками. Это повлекло за собой развитие задних конечностей, которые постепенно удлинились и стали более мощными, чем передние, и у них должны были развиваться цепляющие функции. Когти на пальцах задних ног, поскольку они не были втягивающимися, как у кошачьих, становились менее полезными: постепенно укорачиваясь и уплощаясь, они превратились в ногти. На пальцах передних конечностей у таких насекомоядных сохранились когти, способствуя фиксации за ту ветку, на которую совершается прыжок. Подобного рода специализация конечностей отмечается у примитивного примата анагале гобийского (Anagale gobiensis) из Олигоценовых отложений Монголии: его считают родственником тупайи.
Вторая группа древнейших приматов выделилась из долгопятовых.
У других древних насекомоядных специализация в прыжках помимо соответствующего изменения задних конечностей в целом вызвала также и удлинение пяточного отдела стопы, так что их задние конечности стали представлять собой двойной рычаг, позволявшие им, несмотря на небольшие размеры тела, совершать огромные прыжки. В этом случае для фиксации за ту ветвь, на которую зверёк прыгал, одних когтей оказалось недостаточно, и передние конечности должны были развить способность к захвату ветвей. Это привело к удлинению пальцев передней конечности и усилению возможности раздельного манипулирования ими. Необходимость лазания по деревьям привела и к возникновению способности большого пальца кисти противопоставляться остальным, что позволило охватить тонкие ветки. Так сформировались представители подотряда долгопятовых.
Способность держать тело в вертикальном положении развивалась у предков приматов также под воздействием их образа жизни: чем большее значение приобретало лазание по стволам и лианам, тем чаще тело принимало вертикальное положение, а это вело к укреплению позвоночного столба, перестройке мышц спины, изменению функций передних и задних конечностей. Предки долгопятовых чаще, чем предки тупайи, пользовались таким способом лазания, поэтому у них развились вертикальная постановка тела и хватательная способность передних конечностей. Длина морды укорачивалась, а череп становился более округлым, чем у типичных четвероногих животных.
Перескакивание с ветки на ветку и с дерева на дерево вело также к изменению органов зрения. Необходимость точной ориентировки и оценки расстояния при передвижении привела к возникновению стереоскопичности: зрительные оси обоих глаз должны были получить большее схождение, что могло быть достигнуто перемещением глаз вперёд, на лицевую сторону черепа. Долгопятовые отличаются огромными размерами глаз, но эта особенность возникла вследствие ночного образа жизни. В соответствии с прогрессом органов зрения, к которым перешла особо важная роль в передвижении и ориентировке в обстановке, обоняние у приматов по сравнению с наземными животными несколько ослабилось.
Первая группа древнейших приматов, выделившаяся одновременно с долгопятовыми, приспособилась преимущественно к лазанию. Поэтому передние и задние конечности у них специализировались как хватательные. Кисти и стопы их должны были действовать подобно клещам, зажимая ветви или лианы между большими и остальными пальцами. Передвижение прыжками играл второстепенную роль. При лазании вертикальное положение тела было не менее обычным, чем горизонтальное или наклонное, и у потомков третьей группы примитивных приматов с течением времени начали накапливаться черты, всё более и более приближавшиеся к обезьянам. Хватательные способности конечностей привели к образованию специальных приемников осязательных раздражений в виде выпуклых осязательных подушечек, покрытых капиллярными линиями и узорами. У обезьян и человека подобными узорами сплошь покрыта кожа ладоней и подошва ступней. Особая сложность и разнообразие движений в сочетании с сильной подвижностью свойственны большинству древесных млекопитающих, привели к значительному развитию моторных участков мозга. Переход лазающих древнейших приматов, питавшихся насекомыми, к употреблению растительной пищи обогащал их организм более разнообразными веществами. Это тоже стимулировало усложнение и рост объёма мозга, который и у низших приматов развит довольно сильно.

Третья группа животных, переходных от насекомоядных к приматам, дала в начале Палеогена ветвь полуобезьян, или лемуров.
Изменение пищи и увеличение её разнообразия вызвали прежде всего преобразование челюстей и зубов. Часть зубов оказалась лишней и исчезла, другие претерпели изменение формы и структуры, становились способными перетирать любую пищу. Так, если у древних насекомоядных было 44 зуба-столько же, сколько установлено у анагале гобийского, то у ископаемых долгопятовых их количество сокращается до 40-28 (у современных долгопятов-34), а у лемуров-до 40-32.
65-23.5млн.л.н. ПАЛЕОГЕНОВЫЙ Период
63.5-53.5млн.л.н. ПАЛЕОЦЕН
Первая группа (протолемуроиды) - лемуры.
Лемуры известны как из слоёв третичного периода (Палеоцен, Эоцен, Миоцен) Северной Америки, Европы и Азии, так и из четвертичного - о-в Мадагаскар.
Лемуры семейства плезиадапсид, родственного семейству современных мадагаскарских руконожек, известны из Палеоцена и Эоцена Европы и Северной Америки. Их передние зубы (резцы) были очень велики и обладали постоянным ростом.
У семейства адапид резцы стоят более или менее вертикально, невелики, клыки умеренных размеров, зубы стоят без промежутков. Череп адапид низкий и широкий, с сагиттальным и затылочным гребнями, суживается позади орбит, лицевой отдел довольно длинный, глазницы не так велики, как у потомков адапид-мадагаскарских лемуров. Головной мозг был невелик и весьма примитивен. Задние конечности короче, чем у современных лемуров.
Группа адапид явилась основой для развития некоторых мадагаскарских форм лемуров. При развитии типа современных мадагаскарских лемуров подсемейства лемуриновых из типа адаписа произошли следующие изменения: орбиты увеличились, лицевой отдел укоротился, сигиттальный гребень исчез, мозговая коробка сделалась объёмистой, слуховая капсула увеличилась, клык уменьшился в размерах.
Представители подсемейства нотарктиновых известны из Северной Америки. Основная форма - нотарктус из Палеоцена и эоцена; известен почти весь скелет: плечо короткое с надмыщелковым льверстием, бедро длинное без третьего вертела, фаланги пальцев расширены, следовательно, были ногти, большие пальцы кистей и стоп были хватательными. Мадагаскарские ископаемые лемуры относятся к семействам лемурообразных и вымерших археолемуров. Многие из них обладали крупными размерами тела, найдены в торфяных болотах и пещерах в плейстоценовых (ледникового времени) и голоценовых (современных) слоях.
Из лориподобных лемуров недавно стал известен ископаемый представитель - индралори, от которого найден первый М в миоценовых слоях Индии. Индралори является предком для лориевых лемуров Юго-Восточной Азии.
Эоценовые примитивные формы прототарзиоидов (первая группа) дали начало исходным формам обезьян Нового и Старого Света.
53.5-37.5 млн. л. н. ЭОЦЕН
53 млн. л. н. происходят извержения вулканов в районе Срединно-Атлантического хребта.
Вторая группа (прототарзиоиды) - долгопяты.
Долгопят - примитивное ночное животное, ведущее исключительно древесный образ жизни,- прыгает и карабкается по веткам.
Ископаемые долгопяты известны из Палеоцена и Эоцена Северной Африки и Европы, всего 22 рода. Пережитком с тех времён является современный долгопят, к которому из ископаемых близки псевдолори и тетониус; обезьянам Старого Света родственен некролемур. Все ископаемые долгопяты объединяются в семейство анаптоморфид.
Известно 13 родов североамериканских ископаемых форм долгопятовых, из которых один – омомис - обнаружен и в Европе в нижнеЭоценовых слоях Бельгии. В Северной Америке омомис обнаружен в Эоцене Уайоминга. Европейские ископаемые долгопяты представлены несколькими формами.
Псевдолори - крошечный долгопят из Эоцена Франции, известный по остаткам черепа. Основная форма европейских долгопятов - некролемур.
Микрохерус - вид, родственный некролемуру, из Эоцена Англии, Франции и Швейцарии.
Периконодон - вид европейских ископаемых долгопят из Эоцена Швейцарии.
Группа ископаемых долгопятов в своей основной массе пришла в результате специализации органов зрения и конечностей к упадку и в связи с иными причинами биологического порядка вымерла в конце Эоцена. Она дала начало, с одной стороны, современному долгопяту, с другой стороны - группа долгопятов сыграла значительную роль в формировании древнейших форм обезьян.
После дневного сна эти приматы приходят в движение. Они живут на деревьях. У этих приматов острое зрение и невероятно ловкие руки, они помогают им не только перемещаться по веткам, но и собирать любимых насекомых, даже если они находятся под корой. Годиноции-очень находчивые и изобретательные животные. Они живут по одиночке. в ночные часы некоторые Годиноции становятся менее одинокими. Спаривание у этих приматов - часть их обычного ночного ритма. Эоцен Германии.
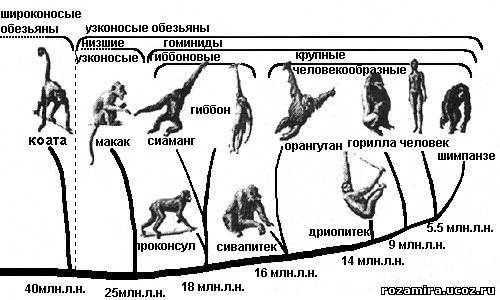
- На рубеже Эоцена и Олигоцена 40-36 млн. л. н. появились широконосые и узконосые обезьяны.
ДОЛГОПЯТЫ (Вторая группа) → узконосые обезьяны.
Узконосые обезьяны развились из древних долгопятов Старого Света типа некролемуров, в середине Эоцена или в его начале.
Апидии - живут хорошо организованными группами и быстро оповещают друг друга при малейших признаках опасности. Разные деревья дают, плоды в разное время, довольно часто им приходится пересекать протоки, а это очень рискованное занятие. Там водятся акулы и крокодилы, а теперь и киты. Прыжки с дерева на дерево это то, в чём Апидиям нет равных. С наступлением прилива некоторые протоки становятся широкими, поэтому Апидиям приходится прибегать к более опасным методам переправы - ближе к воде.
37.5-23.5 млн. л. н. ОЛИГОЦЕН
36.9 млн. л. н. на планету упал метеорит, убивший большинство живого на планете. Вымерло несколько видов морских организмов - радиолярий.
Северный Полюс начал перемещаться в Скандинавию и далее в Карелию (на юго-восток).
36.9 млн. л. н. происходят извержения вулканов в районе Срединно-Атлантического хребта. В конце Палеогена наступило значительное похолодание и одновременно стала развиваться регрессия. Уровень Мирового океана понизился. Многие регионы осушились и превратились в равнинные слабовсхолмлённые области. В значительной мере сократилась ширина экваториального и тропического поясов. Температуры в низких широтах понизились на 5-8 градусов, а в средних и высоких широтах-на 8-12 градусов. Это привело к существенному расширению умеренного пояса и появлению в конце Палеогена и в Неогене отрицательных зимних температур. В горных районах Антарктиды появились первые ледники. С течением времени их толщина и площадь нарастали и возник обширный ледниковый покров.
Началась Лавразийская ледниковая Эра.
Первая группа (протолемуроиды).
Примитивного примата анагале гобийского (Anagale gobiensis) из Олигоценовых отложений Монголии считают родственником тупайи. Череп анагале, длиной 6 см., обладал большим сходством с черепом тупай, но зубная формула очень примитивна. Судя по строению фаланг, на кисти у анагале были когти, а на стопе-ногти.
ВЫСШИЕ ПРИМАТЫ/Широконосые обезьяны - подотряд отряда приматов.
На протяжении всей своей истории был ограничен Южной Америкой.
Первые обезьяны Южной Америки известны с Олигоцена - браниселла из Боливии, долихоцебус и тремацебус из Аргентины и сходны с современными игрункаи и цебусовыми.
ЧЕЛОВЕКООБРАЗНЫЕ ОБЕЗЬЯНЫ
Ископаемые человекообразные обезьяны известны только в Старом Свете, начиная с нижнего Олигоцена.
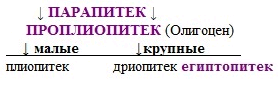
ПАРАПИТЕК - самый примитивный представитель человекообразных обезьян. Найден в Египте (Файюм). Парапитек - исходная форма для позднее развившихся антропоидов, в том числе и человека. Ближайшим потомком его является проплиопитек. ВЫСШИЕ ПРИМАТЫ/ЧЕЛОВЕКООБРАЗНЫЕ ОБЕЗЬЯНЫ
ПАРАПИТЕК → АНТРОПОИДЫ ПАРАПИТЕК → ПРОПЛИОПИТЕК
ПРОПЛИОПИТЕК является предковой формой для всех позднейших человекообразных обезьян. От него в одну сторону пошла линия развития "малых человекообразных обезьян" типа гиббонов; на этой линии одним из промежуточных звеньев является ПЛИОПИТЕК. В другую сторону от проплиопитека пошла линия крупных ископаемых человекообразных обезьян, представленная в Миоцене СИВАПИТЕКОМ, ДРИОПИТЕКОМ и другими формами. Проплиопитек близок к гиббонам. Их сближает, в частности, расположение добавочного бугорка-гипоконулида, который находится точно посередине заднего края талонида. Имеются и отличия от современных гиббонов: челюсть менее крупная, чем у гиббона, первые премоляры развиты слабее. Клыки проплиопитека значительно меньше, чем у гиббона, но крупнее, чем у парапитека. ВЫСШИЕ ПРИМАТЫ/ЧЕЛОВЕКООБРАЗНЫЕ ОБЕЗЬЯНЫ.
ЕГИПТОПИТЕК - очень примитивный дриопитек. Он является предком рамапитека и обладает чертами гоминид. ВЫСШИЕ ПРИМАТЫ/ЧЕЛОВЕКООБРАЗНЫЕ ОБЕЗЬЯНЫ.
ЕГИПТОПИТЕК → РАМАПИТЕК
Эоценовые примитивные формы прототарзиоидов (вторая группа) дали начало исходным формам обезьян Нового и Старого Света.
ДОЛГОПЯТЫ (Вторая группа) → узконосые обезьяны
ВЫСШИЕ ПРИМАТЫ/Низшие узконосые обезьяны (вторая группа) известны в большом числе форм из нижнего Олигоцена, Плиоцена и Плейстоцена Старого Света. Узконосые обезьяны развились из древних долгопятов Старого Света типа некролемуров, вероятно в середине Эоцена или в его начале.
Древнейший их представитель - апидиум (подсемейство мартышковых), известный из нижнего Олигоцена Файюма (Египет) по фрагменту нижней челюсти.
- 25 млн. л. н. мартышкообразные обезьяны: макаки и павианы, отделились от общего ствола предков.
МАРТЫШКИ - самый многочисленный по количеству видов род приматов. Африка.
Мартышки - обезьяны средних размеров, массой тела 3-6 кг и с хвостом (55-100 см.) более длинным, чем голова и туловище (40-65 см.). Самки мельче самцов. Шерсть густая, мягкая и короткая; по окраске она сильно варьирует у разных видов, но зелёные, жёлтые и чёрные цвета преобладают. Для рода характерны яркие локализованные пятна, полосы, усы и борода. Мордочка умеренной длины.
Мартышки встречаются главным образом в дождевых и горных (до 3000 м. над уровнем моря) лесах. Очень быстрые в движениях, они хорошо бегают по земле. Некоторые виды умеют плавать. Спят в густой листве сидя. Питаются листьями, молодыми побегами, плодами, часто совершают набеги на плантации.
Держатся мартышки стадами в 40-50 особей, но состав и размер стада варьирует в различное время дня. Чаще всего в период передвижения в поисках пищи стадо состоит с одним взрослым самцов. У мартышек стада менее организованы, чем у павианов.
МАКАКИ - животные средних и крупных размеров с массой тела 3.5-18 кг, самки значительно меньше. Север Африки, Азия от Пакистана до Японии и Тайваня, от Тибета до Цейлона, на островах Малайского архипелага до Филиппин. Встречаются в тропических дождевых и горных лесах (до 4000 м. над уровнем моря), в умеренных лесах Китая и Японии, а также в травянистых и сухих областях среди кустарников и кактусов в Индии и на Цейлоне.
Макаки - полудревесные, полуназемные животные. Чтобы уберечься от хищников, они спят на деревьях или прячутся в утёсах и скалах. Пища их разнообразна - плоды, корни, молодые листья, насекомые, личинки, моллюски, ракообразные. Они часто совершают набеги на плантации и посевы, где поедают рис, сахарный тростник, кукурузу, картофель.
Макаки живут большими группами, в которые входят более чем один самец и несколько самок с детёнышами разных поколений. В таких группах ярко выражена иерархия. Центральное место занимает сильный самец, затем самки и детёныши. Иногда подчинённые самцы живут изолировано. Стадо занимает определённую территорию в несколько квадратных километров. Часто ареалы обитания отдельных стад перекрывают друг друга, но драки между стадами бывают редко.
Они имеют широкий размах эмоций для общения друг с другом. У них отмечено свыше 30 различных звуков.
ПАВИАНЫ - крупные животные, с массой тела до 25 кг и более. Самки почти в 2 раза легче самцов. У павианов длинная морда (собакоголовые). Клыки сильно развиты. Надбровные валики большие, из-за них глаза смотрят вниз. Ноздри широко открыты и находятся на конце морды. Вся Африка, к югу от Сахары, западная часть полуострова Аравия.
Живут стадами до 80-100 особей. Такие сообщества павианов характеризуются ярко выраженной иерархией. Каждая группа состоит из взрослых особей, подростков и детёнышей. Отношения между членами стада основываются на господстве и подчинении. Господствующее положение принадлежит сильному взрослому самцу. Стадо павианов занимает обычно 2-3 км2. Спят они в расщелинах и под навесами скал, чаще всего сидя, прижавшись друг к другу, детёныши-под защитой матерей. Просыпаются на восходе солнца и начинают активную жизнь в поисках пищи и развлечений.
Павианы всеядны: питаются сочными плодами, молодыми побегами растений, кореньями, ящерицами, насекомыми, яйцами птиц; совершают набеги на поля, сады и огороды.
 ЕДИНСТВО
ЕДИНСТВО